How do organisms evolve new sexual signals when those same signals are needed to start courtship? Zhang et al[1] take advantage of a new form of the Hawaiian field cricket Teleogryllus oceanicus to shed some light on the problem.
Insects utilise a range of different signals for the purpose of communication: chemical, visual, tactile and auditory. Crickets are one of the most iconic in terms of using auditory signals, constructing complex patterns of trills and chirps into songs. In he case of the field cricket, T. oceanicus there are three distinct songs serving separate purposes: aggression, communication, and courtship[2]. Males create these by stridulating (or rubbing together) specialised structures on their wings, so a change in wing morphology therefore has a direct impact on the sound/ pitch of the song and therefore the cricket’s ability to communicate.
Long term monitoring of T. oceanicus on the Hawaiian islands by the research group have revealed exactly this: a new phenotype has arisen in the form of a short-wing morph (fig.1) and the consequences can definitely be heard[3]. Wing length influences the pitch of song produced, resulting in short-wing individuals producing sound of a dramatically different frequency to their counterparts (and in some cases have an inability to make any sound at all. But this isn’t the first time a silent morph has evolved in this species, with a flat-wing morph previously being identified and studied[4]. The short-wing morph has only been detected within the past six years, and has emerged on not just on one, but multiple islands. And with increasing variation in the crickets’ signals, comes the opportunity to study the impact on behaviour, female mate-choice, and potential opportunities for speciation[5].
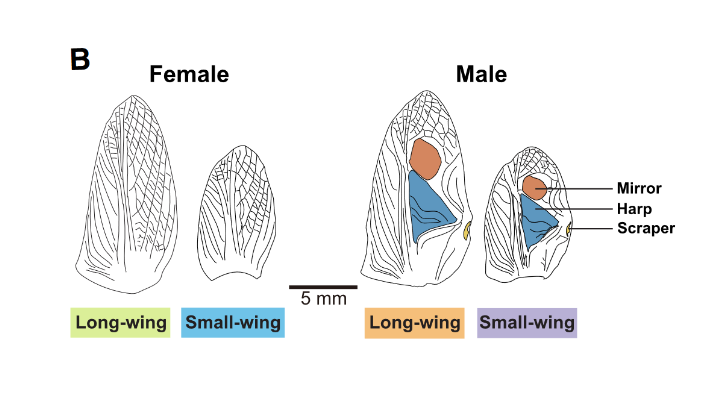
Figure 1: T. oceanicus wing structures. Modified from Zhang et al., 2024
The group decided on the impact of variation in courtship signalling. Changes in these signals can run in tandem with speciation (especially in island systems) as populations which recognise one signal may not recognise a new emerging one, resulting in them becoming isolated through mate-choice. The evolution of new courtship signals can be rare, as if the receiver (in this instance, female crickets) don’t recognise a modified song they can exert selection pressure against the new morph. Courtship in T. oceanicus isn’t a one-step process. Females are initially attracted to the males location by the use of a calling song[6]. When they’re in close enough proximity the males switch to a courtship song and produce chemicals called CHCs (cuticular hydrocarbons). It’s the combination of both chemical and auditory signals by which females make the decision to mount the male in question. To look at the initial difference between the two phenotypes, Zhang et al measured the carrier frequency of the songs, finding that normal males had a range of 4.68 to 5.42kHz. The short-wing males had a significantly wider frequency: 4.85 to 38kHz.
To investigate whether females exhibited a preference as the frequency of courtship song, the group put females in individual containers with a male of one of three types:
normal, short-wing, and short-wing but silent. Mute males were identified as those which attempt to stridulate their wings but could not produce a sound. They then recorded how often females mounted the males, and overall mating success (fig.2).
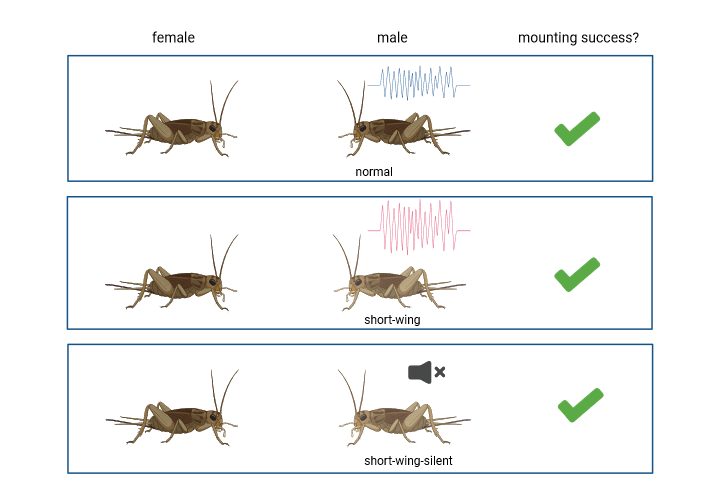
Fig. 2: male phenotypes presented to female T. oceanicus and mating outcome. Created with Biorender.
Surprisingly they found that females mounted males not only regardless of morph, but regardless of if the male was actively courting the female (fig.3). The group interpreted this as ‘female permissiveness’: females not showing a preference and therefore not preventing deviations. This explains how the phenotype has been able to persist in the population: the females don’t adhere to strict guidelines when choosing a mate. Populations which experience bottlenecks (such as those colonising islands) can find that females become more permissive whilst the population size is small. However this doesn’t entirely explain how the mute phenotype has been able to persist outside of the lab, as females need to be able to locate males within an open environment. However the authors do acknowledge a potential issue in the form of sample size. In order to have access to enough short-wing crickets for the study it was necessary to breed more in lab. But there is still quite a small sample size, with which to try and draw conclusions. Another potential factor as to why females may attempt to mate unprompted may lie in chemical signals, ie CHCs. Males that are mute aren’t necessarily unable to produce these, and may have been unintentionally interfering with the behaviour of the females.
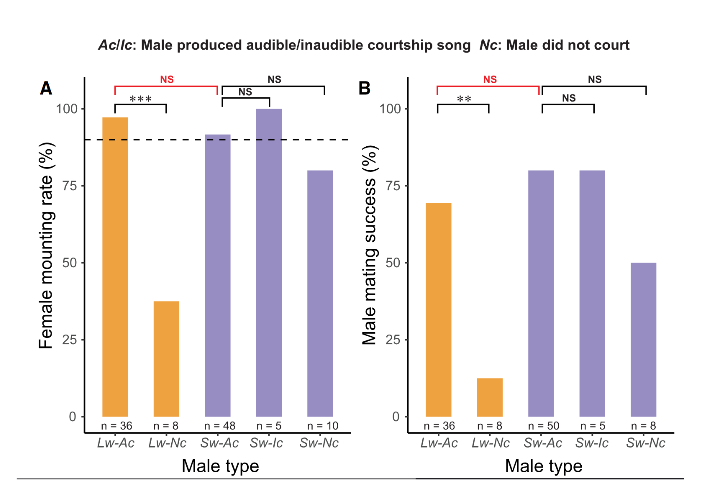
Figure 3: Female acceptance of males. Adapted from Zhang et al., 2024. A: female mounting rate. B: male mating success. Lw-AC: long wing males that produce audible song, Lw-Nc: long wing- males with no courtship sone, Sw-Ac: short-wing males with audible song, Sw-Ic: Short-wing males trying to make song but failing, Sw-Nc: short-wing males with no song.
The findings also raise questions as to why there’s been convergent evolution of why there has been convergent evolution of silent cricket phenotypes on Hawaii. Without this being driven by females, the answer may lie in predation. T. oceanicus is parasitised by a fly: Omis ochracea. This is an eaves dropper, listening in as crickets communicate with each other in order to locate one on which to lay their eggs[7]. In order to distinguish the cricket from other insects in the environment the flies to attuned to the range of frequencies standard crickets produce. Those at the bounds of the higher frequency lie outside of this range, and are less likely to be targeted by the parasitoids. As such, short-wing individuals may actually be at an advantage. This advantage is likely responsible for the morphs rapid emergence and spread across the islands.
Finding organisms where you can watch evolution of signals occurring in real time can be difficult. How do you pick a candidate population and invest resources when you don’t know where and how new signals may evolve? However this research group have the rare opportunity to watch evolution in action in a population that’s already being studied, and potentially have a front-row seat for evolution of a new species of island cricket.
References:
[1] Zhang, R., Rayner, J.G., Bailey, N.W., (2024) Rapid sexual signal diversification is facilitated by permissive females, Current Biology, 34, pp. 403-409, https://doi.org/10.1016/j.cub.2023.11.063
[2] Balakrishnan & Pollack (1996) Recognition of courtship song in the field cricket, Teleogryllus oceanicus, Animal Behaviour, 51 (2), pp. 353-366, https://doi.org/10.1006/anbe.1996.0034
[3] Rayner, J.G., Aldridge, S., Montealegre-Z, F., Baile, N.W., (2019) A silent orchestra: convergent song loss in Hawaiian crickets is repeated, morphologically varied and widespread, Ecology, 100 (8), pp. 1-4, https://www.jstor.org/stable/26749520
[4] Tinghitella, R.M., (2007) Rapid evolutionary change in sexual signal: genetic control of the mutation ‘flatwing’ that renders male field crickets (Teleogryllus oceanicus) mute, Heredity, 100, pp. 261-267, https://doi.org/10.1038/sj.hdy.6801069
[5] Rodriguez, R.L., Ramaswamy, K., Cocroft, R.B., (2006) Evidence that female preferences have shaped male signal evolution in a clade of specialized palnt-feeding insects, Proceedings of the Royal Soceity B, 273 (1601), pp. 2585–2593, http://doi.org/10.1098/rspb.2006.3635
[6] Simmons, L.W., Thomas, M.L., Gray, B., Zuk, M., (2014) Replicated evolutionary divergence in the cuticular hydrocarbon profile of male crickets assocaited with the loss of song in the Hawaiian archipelgo, Journal of Evolutionary Biology, 27 (10), pp., 2249-2257, https://doi.org/10.1111/jeb.12478
[7] Robert, D., Miles, R.N., Hoy, R.R., (1996) Directional hearing by mechanical coupling in the parasitoid fly Ormia ochracea, J. Comp Physiol A, 179, pp., 29-44, https://doi.org/10.1007/BF00193432